4.Фенологическая индикация
В любом ландшафте можно подобрать серию легко и точно регистрируемых сезонных явлений жизни растений и животных, служащих сигналами наступления или приближения практически важных, но трудно регистрируемых моментов годичного цикла природы. Феноиндикаторы–объекты, которые легко и точно отмечают сезонные явления природы. Например, во всей таежной зоне зацветание черемухи сигнализирует о начале пыления елей, а зацветание рябины – о начале пыления сосен. Начало пыления ежи сборной и зацветание таволги свидетельствуют о переходе растительности лугов к массовому цветению; всходы крестоцветных – о близком оживлении листоблошек. По сезонному состоянию некоторых видов растительности можно судить об условиях увлажнения. Зацветание ивы – признак просыхания супесчаной почвы до мягкопластического состояния. Геосистемные феноиндикаторы сигнализируют о наступлении или приближении отдельных этапов годичного цикла всего геокомплекса как целого. Прилет грачей, а в Восточной Сибири – галок, на севере пуночек сигнализирует о приходе весны. Зеленение березы свидетельствует о наступлении разгара весны. По объему информации самыми богатыми являются геосистемные индикаторы, они несут сведения о совокупности основных компонентов. При решении этих проблем индикационная фенология контактирует с биогеграфией и физической географией.
По характеру связи между феноиндикаторами и индуцируемыми ими явлениями следует различать индикаторы прямые и косвенные. Если связь между явлениями причинная, то индикатор прямой. Начало оттаивания водоемов определяет время прилета водоплавающих видов птиц. Начало массового лета насекомых – определяет время прилета насекомоядных птиц. Распускание листьев провоцирует активность листогрызущих насекомых. Часто приходится встречаться с косвенными фенологическими индикаторами, когда индицирующие и индицируемые явления не связаны прямыми причинными связями, но определяются третьим фактором или их группой (например, цветение некоторых видов растений на суше и нерест рыб в соседних водоемах через температурный фактор). Особняком стоит группа сезонных явлений, сроки наступления которых регулируются длиной суточных фотопериодов. Сроки эти настолько устойчивы, что не нуждаются ни в каких феноиндикаторах. Иначе обстоит дело с группой сезонных явлений, регулируемых режимом влажности. Роль влажности резко возрастает в аридных странах и в тропиках. От режима влажности зависят сезонные процессы жизнедеятельности у мхов, сапрофитных и паразитных грибов.
Своеобразна также сезонная динамика жизни теплокровных животных – птиц и млекопитающих. Эти животные в известных пределах сами регулируют свой термический режим. Они более независимы от термического режима геосистемы в целом, но сильнее зависят от режима питания. Наиболее популярны у фенологов наблюдения – весенний прилет и осенний отлет перелетных птиц. Вопрос об индифицировании этих перелетов разработан еще слабо.
Важное значение в фенологии имеет вопрос о территориальных границах применения каждого фенологического индикатора. Некоторые индикаторы сохраняют одно и тоже индикационное значение на всем их ареале, цветение черемухи – знак весны во всем ареале черемухи, цветение липы – разгар полного лета на всей территории. Но иногда индикационная функция в разных частях ареала может меняться. Такие изменения могут проходить по двум причинам: при широком ареале, охватывается несколько природных зон, один и тот же индикатор может иметь разное значение. Лещина в лесной зоне начинает пылить, являясь индикатором начала оживления весны, на юге побережья Кавказа при наличии положительной температуры лещина начинает пылить еще в декабре – январе. Ее роль, как сигнала наступления весны, теряется. С другой стороны – вид – индикатор в разных зонах может быть представлен разными сезонными расами. В лесной зоне тростник зацветает одновременно с созреванием хлебов. В Средней Азии растет другая раса тростника, начинающего цвести лишь 2,5 месяца спустя после созревания хлебов. Такие виды могут быть использованы в качестве зональных и региональных фенологических индикаторов. Встречаются и узколокальные феноиндикаторы. Так, в Комсомольске – на Амуре – начало движения воды в местных ключах сигнализирует о начале субсезона снеготаяния.
Для каждой природной зоны и провинции должны быть составлены свои, местные фенологические индикаторы. В настоящее время имеется система геосистемных фенологических индикаторов для страны, она разработана и изложена в программе массовых фенологических наблюдений.
Фенологические индикаторы могут считаться апробированными, когда точно скоррелированы явления. Местный опыт в подборе фенологических индикаторов без должной проверки переносить в другие географические районы не следует.
3.6 Дендроиндикация и методы её проведения
Наивысшее проявление потенциальных свойств любого растительного организма, популяции, биоценоза, выражающегося в его морфологической структуре, биологических особенностях, продуктивности, в полной мере происходит только в условиях оптимума среды обитания.
Толерантность (устойчивость) вида к комплексу факторов среды обитания заложена в его генотипе.
Есть виды с узкой и широкой амплитудой толерантности, от которых зависит величина ареала вида, варьирование мест произрастания и способность к агрессии, к территориальному захвату, с вытеснением местных видов. Классическим примером с широкой амплитудой толерантности вида в лесостепной и степной зонах России является клён ясенелистный (Acer negundo L), где он в силу своих биоэкологических свойств захватывает самые разные места обитания, вытесняя аборигенные древесные породы — виды тополей, липу мелколистую, вязы, дуб черешчатый, угнетает кустарниковые заросли пойменных лесов, интенсивно внедряется в древостои городских зелёных насаждений.
По современной терминологии этот вид отнесён к эврипотентной группе (с широкой амплитудой толерантности), в отличие от стенопотентной (с узкой амплитудой толерантности).
Различают физиологические — потенциальные диапазоны толерантности видов к природным факторам среды и экологические диапазоны присутствия (экологические потенциалы) — реакции на действие совокупности факторов конкретного района, где ведутся наблюдения. Оба этих показателя определяют, в конечном итоге, индикаторную ценность определённого вида, популяции, ценозов. Поэтому как отдельные виды, так различного типа ценозы по ответной реакции на меняющиеся факторы среды могут служить для её оценки на каждый конкретный момент и на происходящие в растительных группировках изменения при мониторинговых исследованиях. В связи с этим такие виды или ценозы определяются как биоиндикаторы, а применительно конкретно к древесным породам — дендробиоиндикаторы.
Принято выделять два вида дендроиндикации — неспецифическую, когда у вида или ценоза возникают несколько одинаковых экологических реакций на действие отдельных природных факторов, и специфическую, связанную с возникающими изменениями организма или ценоза только с одним каким-либо фактором среды.
Как правило, в природных условиях среды мы сталкиваемся с проявлением неспецифической дендроиндикации. В качестве примера можно привести факт замедления роста, вызываемого несколькими одинаково действующими факторами, находящимися в минимуме, — низкая температура, недостаток влаги, света и др.
Среди видов индикиторов по реакции на действие факторов среды обычно выделяют две группы — чувствительные, когда индикатор резко реагирует отклонениями от нормы жизненных проявлений, и аккумулятивные, когда реакция организма (ценоза) постепенная, медленная, вызванная суммарным накоплением параметров воздействующих факторов и проявляющаяся без резкого нарушения и отклонения от нормы морфофизиологических проявлений у дендроиндикатора или экосистемы.
При постоянном слежении (мониторинге) за объектом дендроиндикации выделяют два его типа — активный и пассивный.
Активный мониторинг связан с исследованиями видимых изменений у организмов под действием экологических факторов на модельных особях видов или ценозах, находящихся в типичных для места мониторинга условиях, и пассивный, когда исследуются отклонения от нормы, а также видимые повреждения на свободно живущих особях или ценозах естественных местообитаний.
Для проведения дендроиндикации учёными разработаны методика и методы наблюдений, которые в зависимости от направления исследований подразделяют на фенологические, морфологические, цитологические, анатомические, физиолого-биохимические, флористические, экосистемные и ряд других (Шульц, 1970, 1981).
Для получения достоверных данных при дендроиндикации необходимо соблюдать следующие четыре требования (Ярмишко В. Т., 1997): получение достаточно точных и воспроизводимых результатов; биоиндикация должна проводиться на возможно большом количестве объектов и с однородными их свойствами; диапазон погрешности при расчётах не должен превышать 20%; методы индикации должны быть нетрудоёмкими, быстрыми и дешёвыми при выполнении.
Среди факторов среды обитания древесных растений в природной или урбанизированной среде важнейшими являются антропогенные, связанные с деятельностью человека, и прежде всего это относится к промышленным и транспортным загрязнениям атмосферы, почвы, вод наземных и грунтовых. Для оценки влияния на древесные породы этой группы факторов разработан целый ряд методов исследования.
Из методов дендроиндикации наиболее широко используется метод фенологической индикации, применительно к древесным растениям определяемый как метод дендрофенологической индикации. Метод основан на выявлении времени наступления определённых фенофаз у растительных организмов, что даёт возможность судить о морфофизиологическом состоянии растений, их реакции на сезонные изменения среды обитания видов и об особенностях самой среды, а это позволяет более грамотно вести технологические мероприятия в лесном и лесопарковом хозяйстве.
Фенологические методы индикации связаны с оценкой хода и учёта нарушений фенологического ритма, выражающегося в сокращении отдельных циклов — длительности роста побегов и вегетации; сокращении обилия в генеративной сфере, репродуктивной способности, изменениями в химизме плодов, всхожести семян, ускорении процессов старения. Эти методы позволяют дать объективную оценку как природной, так и урбанизированной среды обитания растений. Фенологические методы дендроиндикации достаточно полно представлены в работах отечественных ученых — Булыгина, 1982; Николаевского, 1999; Ярмишко,1997; Сергейчик, 1998 и др.
Следующим по значимости методом дендроиндикации является морфобиометрический, изучающий появление у растений под воздействием воздушных и почвенных поллютантов некрозов, хлорозов на коре и листьях, снижение линейного прироста побегов и радиального ствола, снижение облиственности и её площади, длительности функционирования листьев, ускорение отмирания нижних ветвей и, в конечном итоге, сокращение жизни самих растений.
Анатомо-цитологические методы дендроиндикации основаны на выявлении изменений под действием промышленных загрязнителей на уровне клеток и тканей органов растений, выражающихся в нарушениях обменных процессов и, прежде всего, процесса фотосинтеза из-за изменений в эпидермальной, ассимиляционной и проводящей тканях, а также в изменениях, возникающих в хлоропластах, в которых поллютанты концентрируются в наибольшем количестве.
Особой чувствительностью к газообразным поллютантам кислой группы (NO2, SO2, Cl2), соединениям типа (NH3, CH2O и др.) чувствительны биомембраны клеток, которые теряют свойственные им функции регулирования и ограничения проникновения в клетки вышеназванных соединений, что приводит к потере оводнённости и как следствие этого — нарушению фотосинтеза.
Биофизические методы основаны на оценке регистрации pH и rH, электропроводности и электрической ёмкости, биолюминисценции, спектрального анализа исследуемого растительного материала, подвергшегося действию экологических природных и антропогенных факторов, и могут выполняться лишь в динамике. Под действием поллютантов промышленных эмиссий, как показали исследования, у древесных ослабленных растений резко возрастает электросопротивление тканей, увеличиваясь с 80 — 100 до 200 — 500 мОМ (В. Т Ярмишко).
Дендрохронологический метод основан на оценке величины годичных приростов древесины в толщину при анализе годичных колец, реагирующих на изменения тепло- и влагообеспеченности и воздействия промышленного загрязнения в период вегетации. Так, под действием промышленных выбросов двуокиси серы при умеренном воздействии радиальный прирост сосны обыкновенной снижается на 40 — 50%, при сильном — до 80 — 85%. Этот метод достаточно достоверен и прост в применении, позволяет оценить ущерб, нанесённый загрязнением среды лесным экосистемам, и дать ему экономическую оценку (Ярмишко, 1997).
Наиболее чувствительной к загрязнению воздушной среды фтором из трёх древесных хвойных пород — сосны, ели, лиственницы — оказалась ель, проявившая повышенную чувствительность к этому соединению.
Одними из часто применяемых на практике методов индикации являются флористические методы, в основе которых лежат наблюдения и анализ флористической ситуации локальных флор района исследований. При оценке экологической ситуации в сильно загрязнённых районах важно правильно выбрать растительные индикаторы, чутко реагирующие на изменения воздушной среды. Так, в степной зоне Южно-Уральского региона, на Оренбургском газоконденсатном месторождении такими индикаторами являются представители напочвенных лишайников родов Cladonia, Parmelia, Collema, Xantoparmelia, активно реагирующих на загрязнение среды исчезновением в импактной и резким сокращением численности на единице площади в буферной зоне воздействия поллютантов.
В лесных экосистемах при возрастании в воздухе предельно допустимых концентраций загрязняющих веществ гибнут в первую очередь самые чувствительные виды лишайников, относящихся к родам Usnea, Bryopogon, Alektoria; при дальнейшем увеличении в воздухе токсических веществ исчезают и более устойчивые виды лишайников родов Parmelia, Hypogymnia.
Наиболее чувствительны из древесных пород к действию поллютантов хвойные: пихта, сосна, ель. Лиственные древесные повреждаются в меньшей степени. Их гибель обычно связывается с очень высоким многолетним воздействием накопительного характера токсических веществ на растительные организмы.
Для оценки состояния, динамики и функций растительных группировок (сообществ), в том числе и лесных экосистем, используют биоценотические методы, сущность которых заключается в оценке биоразнообразия ценозов, подвергшихся и неподвергшихся антропогенному воздействию. Показатель биоразнообразия в этих методах выступает как главный критерий в оценке экологических условий среды и является индикатором адаптации популяций древесных пород в лесных экосистемах отдельных регионов РФ.
Методы биоиндикации, в том числе и дендроиндикации, будут значимы и дадут объективную оценку состояния лесных экосистем только в том случае, если будут соблюдены три комплекса задач:
а) сохранение видов в биоценозах, находящихся под угрозой исчезновения;
б) сохранение видового разнообразия насаждений на определённых территориях;
в) создание особо охраняемых территорий — заповедников, заказников, памятников природы.
Контрольные вопросы Объясните понятия: экологические факторы, экологическая реакция древесных растений, экологические свойства. Что понимают под средой обитания, условиями произрастания и существования растений? Что такое экологическая ниша ботанического вида, норма экологической реакции, экологическая пластичность? Что вкладывается в понятие «область экологической устойчивости вида»? Каковы кардинальные точки экологической устойчивости, экстремальные условия среды обитания? Назовите и охарактеризуйте группы экологических факторов; факторы абиотической и биотической среды.
Древесные растения подкласса астериды
В этот подкласс входят наиболее высокоразвитые группы дву-дольных, характеризующихся многими эволюционно-прогрессив-ными признаками. Число частей цветка небольшое и всегда фикси-рованное, цветки, как правило, сростнолепестные, а семязачатки с одним интегументом. Астериды — преимущественно травянистые растения, часто однолетние.
На долю древесных здесь приходится значительно меньше ви-дов, чем в других подклассах двудольных — около 24 300 из 65 000.
Семейство маслиновые(Oleaceae) — вечнозеленые или листопадные деревья и кустарники с простыми или сложными су-противными листьями. Цветки в кистевидных или метельчатых соцветиях, реже одиночные, обоеполые, ложнообоеполые и типично раздельнополые. Околоцветник четырехчленный, двойной, простой или в связи с переходом к анемофилии полностью редуцированный. Тычинок 2 (3-5), на коротких нитях, пестик из 2 плодолистиков, завязь верхняя (*Ko-4Q _(4)Aq-2-5G0^(2) ). Плоды различных ти-пов: коробочка, костянка, крылатая семянка, ягода. Семейство объединяет до 600 видов 30 родов, произрастающих в тропических, субтропических и умеренных областях.
Род маслина (Olea)-вечнозеленое небольшое дерево, распространенное в тропиках и субтропиках. В Крыму, на Кавказе, в Средней Азии культивируют маслину европейскую, или оливко-вое дерево (О. europaea), интродуцированную из Средиземно-морья. Листья ланцетные или узкоэллиптические, до 10 см дли-ной, кожистые, с загнутыми краями, сверху темно-зеленые, снизу серые. Цветки мелкие, с зеленоватым венчиком. Плод — сочная костянка, внешне напоминающая сливу, черная или зеленая, съедобная, с сильномаслянистой мякотью. Маслина — одно из ценнейших масличных деревьев. Оливковое масло, известное под названием прованского, ценится в пищевой промышленности, ме-дицине, используется в технике.
Род ясень (Fraxinus) -обычно крупные листопадные де-ревья с тонкой, светлой или бурой корой. Почки конические и ша-ровидные, черные или коричневатые. Листья супротивные, непар-ноперистосложные. Цветки в кистевидных или метельчатых много-цветковых соцветиях, образующихся в год, предшествующий цве-тению, в пазушных цветковых или, реже в верхушечных почках. Цветут до распускания листьев или одновременно с облиствением, опыляются ветром. У подавляющей части видов цветки без около-цветника, состоят из 2 тычинок и 1 пестика, часто происходит ре-дукция тычинок или пестика, тогда цветки становятся однополы-ми, а деревья — многодомными и двудомными. Плод — крылатая семянка, созревающая осенью и распространяемая ветром; семя с эндоспермом. Все виды ясеня светолюбивы, требовательны к почве и относятся к породам, образующим широколиственные леса. В роде насчитывается свыше 60 видов, из которых в СССР естественно растут 9 и около 20 интродуцированы.
Ясень обыкновенный (F. excelsior) — стройное полнодревесное дерево до 40 м в высоту и 1 м в диаметре. Кора светло-серая с кра-сивым рисунком неглубоких продольных и поперечных трещин, даже в старости отслаивается с трудом. Почки черные, матовые; побеги толстые, в верхней части слегка сплюснутые, зеленовато-черные, голые. Листья крупные, голые, сверху темно-зеленые, состоят из 7-9, реже 5-15 яйцевидно-ланцетных, заостренных

и пильчатых по краю листочков (боковые — сидячие, верхний -на черешке). Листья распускаются поздно (в период цветения че-ремухи обыкновенной), осенью опадают, обычно не изменяя окраски.
Цветки в метельчатых соцветиях, чаще ложнообоеполые, но могут быть и двуполыми. Преобладают смешаннополые плодоно-сящие деревья, но встречаются и мужские. Зацветает ясень одно-временно с распусканием ростовых почек. У него имеются рано-, позднораспускающаяся и промежуточная фенологические формы.
Плод — продолговатая крылатая семянка с бурым, винтообразно изогнутым крылом (рис. 81). Масса 1000 шт. 60-72 г. Созревают плоды осенью и опадают в осенне-зимний период. Для семян характерен продолжительный период покоя, возрастающий от юж-ных границ ареала к северным. Поэтому семена требуют перед посевом стратификации; семядоли всходов ланцетные, первые листья простые, узкоэллиптические.
Растет ясень быстро, живет 250 лет и более, плодоносить на-чинает при одиночном произрастании с 15-20 лет, в насажде-ниях — с 30-40 лет. Хорошо возобновляется порослью от пня. Растет в широколиственных лесах европейской части СССР, Кры-ма, Кавказа. Чистые ясеневники образует редко, обычно встреча-ется в смеси с другими широколиственными породами.
Древесина ясеня ценится в авиастроении, мебельном производ-стве при изготовлении особо красивого фанерного шпона. Кора идет на дубление кож и используется в медицине. Ясень — краси-вое аллейное и парковое дерево.
Ясень маньчжурский (F. mandshurica) — вид, замещающий ясень обыкновенный на Дальнем Востоке. Растет в хвойно-широ-колиственных лесах, обычно по долинам рек, нередко вместе с в. японским. От я. обыкновенного отличается тупочетырехгран-ными побегами, наличием коротких черешков у боковых листочков листа, полной двудомностью.
Ясень узколистный (F. angustifolia) и ясень сирийский (F. sy-riaca) — близкие к я. обыкновенному среднеазиатские виды. Часто встречаются в озеленении городов республик Средней Азии. У я. узколистного листья из (2) 3-5 пар узколанцетных листочков, зубчатых до нижней четверти; листья я. сирийского состоят из 1-2 (3) пар листочков, зубчатых только в верхней половине.
Ясень цветочный, или манновый (F. ornus) — дерево широко-лиственных лесов Закавказья и Карпат. Его цветки имеют хорошо выраженный околоцветник из белых, сросшихся у основания лепе-стков. Цветет одновременно с облиствением. Листья из 7 эллип-тических листочков. Ценен как источник манны — сладковатого, застывающего на воздухе сока, вытекающего при подсочке вет-вей. Манну применяют в медицине. Красивое парковое дерево, более теплолюбивое и менее зимостойкое, чем я. обыкновенный.
Ясень пенсильванский, или пушистый (F. pennsylvanica) -дерево 20-23 м высотой и свыше 0,5 м в диаметре. Растет по до-линам рек и берегам озер в Северной Америке. Побеги серые, опу-шенные; почки ржаво-бурые; листья из 5-9 черешчатых яйце-видно-эллиптических или широкоэллиптических светло-зеленых и волосистых снизу листочков. Осенью желтеют и опадают раньше, чем я. обыкновенного. Двудомен, цветет до облиствения. Крылатка плода прямая, продолговато-эллиптическая, охватывает только верхнюю часть веретенообразного плодика (рис. 82). Растет бы-стро, живет до 100-150 лет. Светолюбив, зимостоек, к почве сред-

нетребователен. Широко используется в озеленении, особенно в го-родах таежной зоны СССР.
Ясень ланцетный, или зеленый (P. lanceolata) — североамери-канский, близкий к предыдущему виду. От я. пенсильванского от-личается голыми побегами, ярко-зелеными, голыми, неопушен-ными листьями с сидячими ланцетными, по краю острозубчатыми листочками. Крылатки мелкие, до 4 см длиной, крыло обратнолан-цетное, охватывает плодик наполовину (см. рис. 81 и 82). По зимо-стойкости этот ясень почти не уступает пушистому, но значительно более засухоустойчив (ксерофит). Широко применяется в степном лесоразведении и в озеленении городов лесостепной и степной зон СССР.
Род бирючина (Ligustrum) — вечнозеленые или листопад-ные небольшие деревья и кустарники. Листья простые, цельно-крайние. Цветки обоеполые, четвертого типа, с чашечкой и белым венчиком, собраны в конечные, прямостоячие кисти или метелки. Плод — малосеменная черная ягода, семена трехгранные. Цветет после облиствения — в начале — середине лета, опыляется насе-комыми (медонос), плоды созревают осенью.
В Западной Украине, Молдавии, Крыму и на Кавказе в кустар-никовых зарослях растет бирючина обыкновенная (L. vulgare) —
полувечнозеленый кустарник 2-3 м высотой. Листья ланцетные; кожистые. Размножается семенами, корневыми отпрысками, от-водками и порослью от пня.
Широко используется в озеленении.
Род сирень (Syringa) -листопадные кустарники, реже де-ревья, с простыми, супротивными, черешчатыми, цельнокрайними листьями (рис. 83). Цветки обоеполые, с чашечкой и венчиком (*К(4)С(4)А20^2), белые или окрашенные в иные тона, собраны в крупные, конечные или пазушные пирамидальные метелки. Плод — двухгнездная, двухстворчатая коробочка с 4 кожистыми крылатыми семенами. Цветут сирени после облиствения, опы-ляются насекомыми (медоносы), семена созревают и опадают осенью.
В СССР наиболее распространена сирень обыкновенная (S. vulgaris) — кустарник до 5 м высотой родом с Балканского полу-острова. Побеги серые, завершаются парой супротивно располо-женных овальных почек. Листья округло-треугольные, гладкие, осенью обычно опадают зелеными. Цветет первой из сиреней, в конце весны — начале лета. Цветки фиолетовые, розовые или белые, душистые. У садовых сортов, которых известно свыше 500, окраска цветков может быть иной, а цветут они на 2-4 недели позже. Сирень растет быстро, неприхотлива к почвенным усло-виям, очень зимостойка. Чрезвычайно популярна в озеленении и в СССР разводится повсеместно. Культурные сорта менее зимо-стойки. Размножают сирень семенами, черенками, отводками. Она способна давать обильные корневые отпрыски и может служить хорошим закрепителем почв.
В практике озеленения нередко этот вид путают с с. персид-ской (S. х persica), которая отличается от с. обыкновенной лан-цетно-заостренными листьями, рыхлыми метельчатыми соцвети-ями, более поздним цветением и сильным специфическим запа-хом цветков. По зимостойкости с. персидская уступает с. обыкно-венной и поэтому может применяться в озеленении только в рай-онах со сравнительно мягким климатом (Прибалтика, средняя и южная части европейской территории СССР, Кавказ и др.). Пред-положительно родина этого вида — Китай.
Сирень венгерская, или восточно-карпатская (S. josikaea) -кустарник или дерево до 8 м высотой. От с. обыкновенной отли-чается наличием на старых ветвях белых чечевичек, на концах побегов — яйцевидной конечной почки, более крупными, заост-ренными с обоих концов, слегка морщинистыми по жилкам листь-ями, которые осенью принимают фиолетово-бурую окраску. Цвет-ки розово-фиолетовые, в конечных и пазушных метелках. Зацве-тание с. венгерской служит индикатором наступления фенологи-ческого лета. Размножается семенами. Широко используется в озе-ленении. Сирень венгерская — редкий эндемик Карпат, в СССР за-несена в «Красную книгу СССР».

Сирень мохнатая (S. villosa) — внешне схожа с с. венгерской, которую замещает в широколиственных лесах Дальнего Востока. Отличается от последней опушением листьев. Зимостойка.
Сирень амурская, или трескун (S. amurensis) -дерево до 20 м в высоту и 30 см в диаметре ствола. Листья эллиптические или овальные, длиннозаостренные, плотные, гладкие, осенью жел-теют и опадают раньше, чем у других видов сирени. Цветки мел-кие, белые, собраны в крупные (до 30 см) метелки, цветут в пер-вой половине лета. Возобновляется семенами и порослью от пня, растет быстро, живет свыше 100 лет. Зимостойка. Распространена в широколиственных лесах Дальнего Востока. Широко исполь-зуется в озеленении. Древесина идет на разные поделки и изготов-ление лыж.
Семейство ж и м о л о ст н ы е (Caprifoliaceae) включает до 500 видов, относящихся к 15 родам листопадных или вечнозеленых кустарников (иногда вьющихся), невысоких деревьев или трав. Распространены жимолостные главным образом в Северном полу-шарии, в умеренном и субтропическом поясах, кроме пустынь. Листья супротивные, цельные и лопастные (калина), реже пе-ристо- или тройчатосложные (бузина). Цветки обычно с прицвет-никами и прицветничками, в сложных верхоцветных метельчатых, щитковидных полузонтиках или одиночные. Цветут после облист-вения, реже до него. Опыляются насекомыми, многие являются хорошими медоносами. Цветки актиноморфные или зигоморфные, чашечка с короткой трубкой, срастающейся с завязью; венчик трубчатый, колокольчатый, воронковидный, белый или иной яркой окраски; тычинок 2-5, прикрепленных к трубке венчика; пестик из 2-5 плодолистиков; завязь нижняя и полунижняя, 1-5-гнезд-ная, с одним или многими семязачатками в каждом гнезде <*/ К(з_5)С(з_ 5)A.(2_5)G(3_9)-)- Плод ягодовидный, костянковид-ный или коробчатый, раскрывающийся, реже нераскрываю-щийся. Семена с эндоспермом.
Род бузина (Sambucus) — листопадные, небольшие деревья, кустарники, иногда травы. Почки крупные, побеги с рыхлой серд-цевиной. Листья непарноперистые или тройчатые; цветки мелкие, белые или желтоватые, в конечных зонтиках или метелках, цветут после облиствения. Плоды сочные, костянковидные, с 3-5 косточ-ками (рис. 84).
Бузина черная (S. nigra) -дерево до 10 м высотой. Листья из 5-7 (3-9) яйцевидно-эллиптических, пильчатых по краю лис-точков. Цветки белые, в конечных, прямостоячих зонтиковидных метелках, до 20 см в диаметре. Цветет в первой половине — сере-дине лета. Плоды блестяще-черные, до 8 мм в диаметре, съедобные, созревают в конце лета. Естественно растет от Прибалтики до низовий Дона, в Крыму и на Кавказе.
Используется в озеленении; цветки, плоды, листья, кора и кор-ни применяются в медицине.
Бузина кистистая, или красная (S. racemosa) — кустарник или дерево до 5 м высотой. Отличается крупными цветковыми поч-ками, распускающимися очень рано, почти одновременно с рябин-ником, продолговато-яйцевидными, длиннозаостренными листоч-ками листьев, желтовато-белыми цветками, собранными в яцевид-но-конические метелки. Цветет в разгар весны, одновременно с че-ремухой обыкновенной. Плоды красные, мелкие, созревают в сере-дине лета, несъедобны. Естественно растет в Верхнем Днестровье, но в культуре распространена очень широко — от средней тайги до Кавказа, где во многих местах стала обычным компонентом дендрофлоры.
Используется в озеленении, лесных культурах, в медицине.

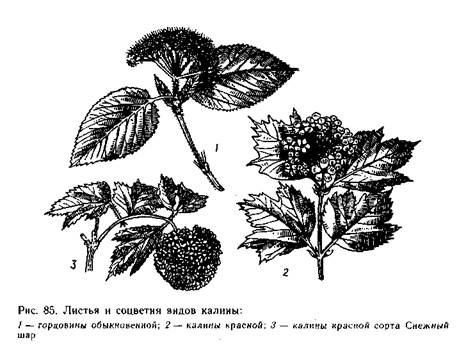
Род калина и гордовина (Viburnum) -листопадные и вечнозеленые кустарники или небольшие деревья с простыми цель-ными или пальчато-лопастными листьями. Соцветия простые или сложные, зонтиковидные, щитки или метелки (рис, 85). Цветки белые или розовые, правильные, мелкие; у многих видов соцветие окаймлено крупными, зигоморфными стерильными цветками, слу-жащими для привлечения насекомых-опылителей. Плод сочный, костянковидный, с одной косточкой, обычно сжатой с боков. Цве-тет калина в конце весны — начале лета, плоды созревают осенью.
Ниже характеризуются 3 листопадных вида.
Калина обыкновенная, или красная (V. opulus) — обычно кустовидное, небольшое дерево с пальчато-лопастными листьями, с крупными стерильными краевыми цветками и мелкими плоду-щими. Костянки красные. Растет в северных лесах в подлеске, по опушкам, на плодородных влажных почвах. На Дальнем Восто-
ке к. обыкновенную замещает весьма схожая с ней калина Сард-жента (V. sargentii). Отличается довольно мелкими краевыми сте-рильными цветками.
Гордовина обыкновенная (V. lantana) отличается густым че-шуйчато-звездчатым опушением побегов, почек и листьев, голыми (без чешуй) почками — узкими вегетативными и крупными шаро-видными цветочными; продолговато-яйцевидными или эллиптиче-скими, морщинистыми листьями; соцветия без краевых стерильных цветков. Незрелые плоды красные, зрелые — черные, съедобные. Растет на юге европейской части СССР и на Кавказе.
Калины широко применяют в озеленении. У к. обыкновенной особенно ценится садовый сорт Снежный Шар (V. opulus ‘Ro-seum’) — с крупными шаровидными соцветиями, состоящими из одних белых стерильных цветков. По зимостойкости значительно уступает основному виду.
Род жимолость (Lonicera) — листопадные и вечнозеленые прямостоячие или вьющиеся кустарники. Листья простые, обычно цельные, иногда сросшиеся основаниями (рис. 86). Цветки сидят попарно в пазухах листьев или собраны в мутовки; зигоморфные, двугубые, верхняя губа состоит из четырех лепестков, нижняя -из одного. Плоды ягодовидные, с остатками чашечки на вершине; парные завязи нередко срастаются, образуя соплодие как бы с об-щей завязью. Размножаются жимолости семенами, дают поросль

от пня. Все они хорошие медоносы, многие применяются в озеле-нении и медицине.
Растут жимолости в подлеске лесов, по опушкам, образуют кустарниковые заросли. Всего в роде насчитывается свыше 180 видов, из которых в СССР естественно растет 61 и около 100 интро-дуцировано. Ниже характеризуются 7 листопадных видов этого рода.
Жимолость Палласа (L. pallasii) -кустарник до 1,5 м высо-той с красновато-бурыми побегами. Листья продолговато-ланцет-но-эллиптические, до 5 см длиной, темно-зеленые. Цветки парные, в пазухах листьев, поникают на цветоносах, кремово-желтоватые, цветут вскоре после начала облиствения. Соплодия овальные, до 12 мм длиной, черно-синие, горькие, созревают в первой полови-не лета и быстро опадают. Растет по таежным лесам в европейской части СССР, в Западной и Восточной Сибири, доходя на север почти до Полярного Круга.
Близкими к этому виду являются жимолость синяя (L. caeru-lea), растущая в Карпатах, и жимолость балтийская (L. baltica), естественно растущая на северо-западе СССР. Все эти 3 вида широко применяются в озеленении под общим названием жимо-лость голубая, или синяя.
Жимолость съедобная (L. edulis) — кустарник, внешне схожий с ж. Палласа. Отличается съедобными плодами, по вкусу напоми-нающими голубику. Растет в Восточной Сибири и на Дальнем Востоке. В культуре используется до Кольского полуострова.
Заслуживает значительно более широкого разведения как ценный плодовый, лекарственный и декоративный кустарник.
Жимолость обыкновенная, или лесная (L. xylosteum) -кустарник 1-3 м высотой с серыми побегами. Листья широко-эллиптические, острые или притупленные, до 6 см длиной, барха-тисто-опушенные или сверху голые (рис. 86). Цветки парные, пазушные, прямостоячие, желтовато-белые, цветут в конце весны. Плоды парные, свободные или сросшиеся только основаниями, созревают во второй половине лета. Растет в лесной части евро-пейской территории СССР и в Западной Сибири, заходя на восток до Алтая. Ценный вид для озеленения.
Жимолость татарская (L. tatarica) — крупный кустарник 4-6 м высотой с желтовато-бурыми побегами. Листья яйцевидно-ланцетные, заостренные или туповатые, с закругленным или почти усеченным основанием, до б см длиной и 3 см шириной, голые. Цветки крупные, парные, пазушные, прямостоячие, от белых до темно-розовых, цветут в конце весны — начале лета. Плоды пар-ные, свободные или слабосросшиеся, шаровидные, до 7 мм в диа-метре, красные, оранжевые или желтые, созревают в середине лета.
Естественно растет в лесостепи и степи, по поймам рек в полу-пустыне и пустыне — от Средней Волги и Южного Урала до Байкала, на юг — до центральноазиатских предгорий. Зимостойка и засухоустойчива. Широко используется в озеленении и в поле-защитном лесоразведении.
Жимолость каприфоль козья (L. caprifolium) — вьющийся кустарник с побегами, достигающими 3 м длиной. Листья эллипти-ческие, до 10 см длины, сверху темно-зеленые, снизу сизоватые. Верхние 2-3 лары листьев срастаются в эллиптический диск (рис. 87). Цветки собраны в 1-2 мутовки в пазухах сросшихся листьев; венчик 4-5 см длиной, от беловатого до темно-розового, позже желтеющий. Плоды свободные, оранжево- красные. Цветет в начале лета, плоды созревают во второй половине — в конце лета. Размножается семенами и вегетативно. Растет на Кавказе, часто используется для вертикального озеленения в различных районах СССР. Зимостойка, неприхотлива.
Род снежноягодник (Symphoricarpos) — листопадные невысокие кустарники с простыми, короткочерешчатыми, цельно-крайними или неправильно-выемчато-зубчатыми листьями. Цветки мелкие, колокольчатые, розовые или белые, в пазушных или конеч-ных кистевидных соцветиях. Плоды белые, шарообразные, ягодо-видные, обычно в кистевидных гроздьях. Снежноягодники цветут с первой половины лета до морозов. В конце лета и осенью на побе-гах можно видеть цветки в разных фазах развития, а также зало-жившиеся, формирующиеся и созревшие плоды (рис. 87). Такой тип биологического ритма снежноягодника свидетельствует о том, что его предками были тропические растения.

В озеленении городов и населенных мест СССР наиболее часто встречается североамериканский вид — снежноягодник белый (S. albus).
Род вейгела (Weigela) -листопадные кустарники 1,5-2 м высотой, с почти сидячими простыми, яйцевидно-эллиптическими или ланцетными, по краю пильчатыми листьями. Цветки одиноч-ные или по нескольку в пазухах верхних листьев, крупные коло-кольчатые (рис. 88), розовые, белые, желтые или темно-красные, цветут с конца весны до середины лета. Плоды — цилиндрические коробочки, раскрывающиеся двумя створками. Семена мелкие, обычно крылатые. Размножается семенами и черенками.
Вейгелы ценятся в озеленении как красиво- и длительноцве-тущие кустарники. В СССР естественно распространены в При-морье Дальнего Востока вейгела ранняя (W. ргаесох) и вейгела Миддендорфа (W. middendorfiana). Первый вид цветет в мае -июне пурпурово-розовыми цветками; второй — в мае — июле цветками бледно-желтого цвета.
1. Каковы наиболее характерные признаки отличия древесных растений отдела покрытосеменные от отдела голосеменные? Какие классы и подклассы включает отдел покрытосеменные; каково общее число семейств, родов и видов покрытосеменных растений?
2. Какими биологическими особенностями характеризуются растения классов двудольные и однодольные? Каковы роль древесных растений этих классов в обра-зовании древесной растительности Земли и СССР; их хозяйственное значение?
3. В каких природных зонах и горных районах нашей страны произрастает наибольшее число видов лиственных древесных растений, занесенных в «Красную книгу СССР»? Какие редкие и исчезающие виды деревьев, кустарников и лиан флоры СССР (по регионам) вы знаете?
4. Каковы характерные особенности семейств магнолиевые *, лимонниковые и лавровые? Диагностические признаки различия входящих в эти семейства изуча-емых родов и видов, их географическое распространение, роль в образовании древесной растительности СССР и хозяйственное значение.
5. Какие биологические особенности характерны для семейства барбарисовые и входящих в него родов барбарис и магония? Какие виды этих родов вы знаете; каковы их роль в образовании древесной растительности СССР, хозяйственное значение?
6. Каковы отличительные черты семейств гамамелисовые, платановые, сам-шитовые и тутовые, характерные диагностические признаки входящих в эти се-мейства родов и видов, их роль в образовании лесов СССР и хозяйственное значе-ние? Какие виды деревьев указанных семейств и по каким районам нашей страны занесены в «Красную книгу СССР»?
7. Какими особенностями характеризуются семейства ильмовые и каркасо-вые; какие роды и виды относятся к этим семействам, чем эти виды и роды отли-чаются друг от друга; какова их роль в образовании лесов СССР, в чем состоит хозяйственное значение?
8. Какова общая характеристика семейств буковые, березовые, лещиновые, ореховые? Морфолого-биологическис особенности и диагностические признаки отличия родов бук, орех, каштан, береза, лещина, хмелеграб, орех и лапина, их важнейшие представители во флоре СССР, хозяйственное значение. Какие виды дуба, березы, лещины, хмелеграба и лапины по районам нашей страны занесены в «Красную книгу СССР»?
9. Какие виды семейств березовые, ильмовые, буковые и ореховые явля-ются главнейшими образователями групп мелколиственных и широколиственных формаций лесов СССР? Морфолого-биологические особенности, диагностические признаки различия этих видов деревьев, их экологические свойства, ареалы, хозяйственное значение.
10. Каковы характерные особенности семейств маревые и гречишные? Диаг-ностические признаки различия входящих в эти семейства родов саксаул, солянка и джузгун, изучаемые представители указанных родов, их роль в образовании древесной растительности СССР, хозяйственное значение.
11. Какими морфолого-биологическими особенностями характерен род тама-рикс, к какому семейству он относится; каковы его изучаемые представители, их роль в образовании древесной растительности СССР и хозяйственное исполь-зование?
12. Каковы общая характеристика семейства ивовые, роды этого семейства, изучаемые виды, диагностические признаки их различия, роль в образовании лесов и кустарниковых зарослей СССР, хозяйственное использование?
13. Какими морфолого-биологическими особенностями и экологическими свойствами характеризуются род тополь и его подроды? Важнейшие виды тополей во флоре СССР, их хозяйственное значение. Какова дендрологическая характе-
* При изучении древесных растений студент обязан знать латинские назва-ния семейств, родов и видов.
ристика осины как важнейшего образователя группы мелколиственных формаций лесов СССР?
14. Каковы характерные особенности семейств актинидиевые, вересковые и волчниковые, изучаемые роды и виды этих семейств, их роль в образовании дре-весной растительности СССР, хозяйственное использование? Какие виды семейств вересковые и волчниковые занесены в «Красную книгу СССР*?
15. Каковы систематическое положение рода липа, его морфолого-биологи-ческие особенности и экологические свойства, важнейшие представители в лесах СССР, хозяйственное значение?
16. Каковы характерные особенности семейств гортензиевые и крыжовнико-вые; какие роды и виды относятся к этим семействам; каковы их диагностические признаки, различия, роль в образовании растительного покрова СССР, хозяй-ственное значение?
17. Какими морфолого-биологическими особенностями характеризуются че-тыре подсемейства розоцветных: спирейные, розовые, яблоневые и сливовые. Назо-вите изученные роды и виды этих подсемейств. Каковы диагностические признаки их различия, роль в образовании растительного покрова СССР, хозяй-ственное использование?
18. Каковы характерные особенности семейств мимозовые, цезальпиновые и бобовые, изучаемые роды и виды этих семейств, диагностические признаки их различия, роль в образовании растительного покрова нашей страны, хозяйственное использование?
19. Какие виды семейств гортензиевые, крыжовниковые, розоцветные, мимо-зовые, цезальпиновые, бобовые и по каким регионам нашей страны занесены в «Красную книгу СССР»?
20. Какими морфолого-биологическими особенностями характеризуются роды эвкалипт, феллодендрон, айлант, фисташка, сумах и скумпия? К каким семей-ствам относятся эти роды; изучаемые виды родов, диагностические признаки их различия, районы естественного произрастания или искусственного разведения в СССР, хозяйственное значение?
21. Какими характерными признаками обладает род клен; каковы его систе-матическое положение, изучаемые виды, диагностические признаки их различия, роль в образовании древесной растительности СССР, хозяйственное значение? Какими морфолого-биологическими особенностями и экологическими свойствами обладают представители рода; каков ареал клена остролистного как важнейшего образователя группы широколиственных формаций лесов СССР?
22. Каковы характерные особенности семейств кизиловые, аралиевые, кру-шиновые, виноградовые и лоховые? Какие изучаемые роды и виды относятся к этим семействам; каковы диагностические признаки их различия, роль в обра-зовании растительного покрова СССР, использование в народном хозяйстве? Есть ли среди растений указанных семейств редкие и исчезающие виды, занесен-ные в «Красную книгу СССР»?
23. К каким семействам относятся каштан посевной и конский каштан обык-новенный; каковы диагностические признаки различия этих видов деревьев, их роль в образовании древесной растительности СССР, использование в народном хозяйстве?
24. Каковы характерные черты семейства маслиновые; какие роды и виды семейства вы знаете? Каковы диагностические признаки их различия, роль в обра-зовании древесной растительности СССР, хозяйственное значение? Какова денд-рологическая характеристика ясеня обыкновенного как важнейшего образователя группы широколиственных формаций лесов СССР? Каких представителей семей-ства маслиновые, занесенных в «Красную книгу СССР», вы знаете? Где нахо-дятся районы их охраны в нашей стране?
25. Какова общая характеристика семейства жимолостные? Каковы морфо-лого-биологические особенности входящих в него родов бузина, калина, жимо-лость, снежноягодник и вейгела? Какие виды этих родов вы изучили? В чем состоят диагностические признаки их различия, роль в образовании древесной растительности СССР и хозяйственное использование? Какие виды семейства жимолостные занесены в «Красную книгу СССР»?
26. Какими диагностическими признаками различия характеризуются важ-нейшие образователи групп мелколиственных (осина, березы повислая, пушистая, плосколистная и даурская, ольха черная) и широколиственных формаций лесов СССР (буки лесной, восточный, дубы черешчатый, скальный, монгольский, орех грецкий, липа мелколистная, вязы гладкий, голый, клен остролистный, ясень обыкновенный)? К каким семействам относятся эти лесообразователи; каковы их биологические особенности, экологические свойства и ареалы?
27. Какие древесные растения со съедобными плодами вы знаете? К каким семействам относятся эти виды; каковы их жизненные формы и в каких природных зонах (горных странах) СССР они произрастают?
28. Какие виды древесных растений применяют в степном и полезащитном лесоразведении и для закрепления подвижных песков? К каким семействам отно-сятся эти виды; каковы их жизненные формы, биологические особенности и эко-логические свойства?
29. Какие лиственные деревья и кустарники, интродуцированные в СССР, наиболее широко применяют в лесном хозяйстве, степном и полезащитном лесо-разведении? К каким семействам относятся эти виды и из каких стран они интро-дуцированы?
ПРИЛОЖЕНИЯ
Приложение 1. Природные зоны и горные страны СССР
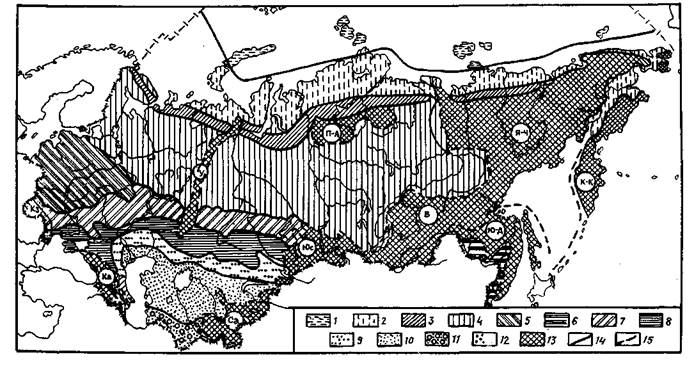
Условные обозначения природных зон:
1 -ледяная; 2 — тундра; 3 — лесотундра; 4 — тайга; 5 — смешанные леса Русской рав-нины; 6 — муссонные смешанные леса Дальнего Востока; 7 — лесостепь; 8 — степь; 9 -полупустыня; 10 — пустыня умеренного пояса; 11 — субтропическая пустыня; 12 — среди-земноморская зона; 13 — горные страны с вертикальной зональностью; границы: 14 природных зон; 15 — горных стран; горные страны: К — Карпаты; У — Урал; K3 — Кав-каз; Са — Средне-азиатская горная страна; Юс — Южносибирская; П-А — Путорано-Анабарская; Б — Байкальская; Ю-Д — Южно-Дальневосточная; Я-Ч — Якутско-Чукот-ская; К-К-Камчатско-Курильская
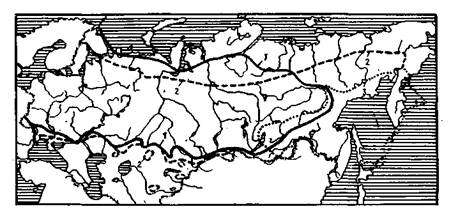
Приложение 2. Ареалы главнейших лесообразующих пород СССР
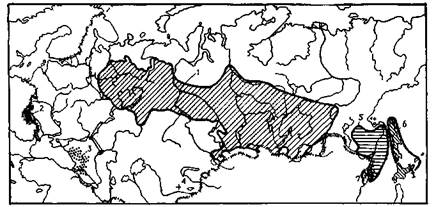
2.1. Ареалы видов пихты:
1 — сибирской; 2 — белой; 3 — кавказской; 4 — Семенова; 5 — бслокорой; 6 — сахалин-ской; 7 — цельнолистной

2.2. Ареалы видов ели:
1 — европейской; 2 сибирской; 3 — восточной (кавказской); 4 — Шренка; 5 — аян-ской

2.3. Ареалы видов лиственницы:
1 — сибирской; 2 — Гмелина (даурской); 3 — Чекановского

2.4. Ареалы видов сосны:
1 — обыкновенной; 2— кедровой сибирской (кедра сибирского); 3 — кедровой стлани-
ковой (кедрового стланца)
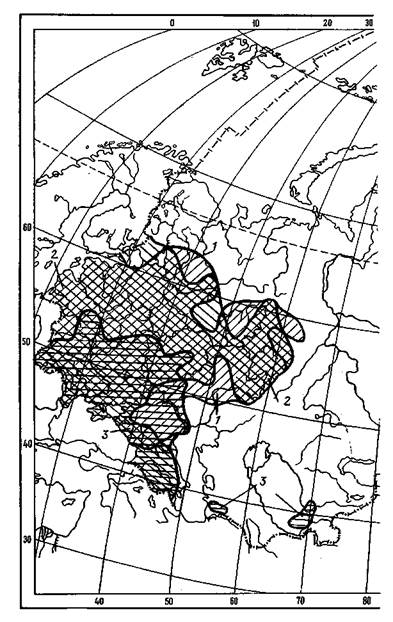
2.5. Ареалы видов вяза:
1 — гладкого; 2 — голого (шершавого, или ильма горного); 3 — граболистного (поле-вого)

2.6. Ареалы видов лиственных деревьев:
1 — осины (тополя дрожащего); 2 — ольхи черной; 3 — березы даурской; 4 — березы
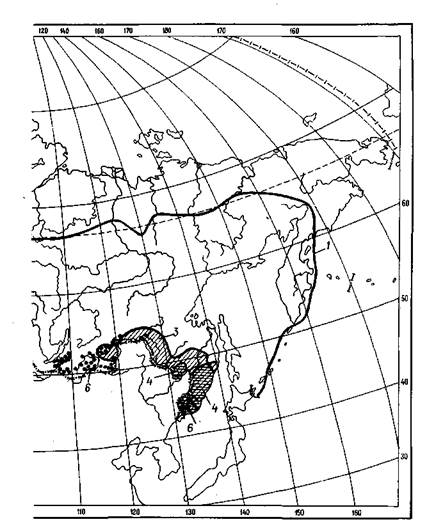
ребристой; 5 — ореха грецкого; 6 — вяза мелколистного (приземистого)
 |
2.7. Ареалы видовберезы:
1 — пушистой; 2 — повислой; 3 — плосколистной, шерстистой и каменной (обобщенный ареал)
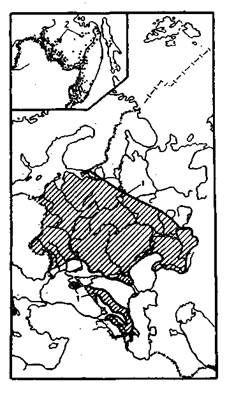 |
2.8. Ареалы видов дуба:
1 — черешчатого; 2 — монгольского

2.9. Ареалы широколиственных видов деревьев:
1 — липы мелколистной; 2 — клена остролистного; 3 — ясеня обыкновенного

Продолжение прил. 3



 |
| Окончание прил.З |
* В алфавитном порядке названий семейств приведен сокращенный перечень редких и исчезающих видов древесных растений флоры СССР. С полным указателем редких видов, их характеристикой, ареалами и рекомендуемыми мерами по сохранению следует знако-миться по официальному изданию «Красной книги СССР» (1984).
Принятые условные обозначения: Д1-4 — дерево соответственно первой-четвер-той величины; K 1-4 — кустарник первой — четвертой величины; Дстл или Кстл — дерево или кустарник стланиковой формы; ПК — полукустарник; Клаз — кустарник лазающий; Л — лиана; Лд — лиана древовидная; Лк — лиана кустарниковая (характеристика упо-мянутых форм и групп роста древесных растений приведена в главе 1); ETC — европей-ская территория СССР.
Система основных дендрофенологических индикаторов для установления факти-ческих сроков наступления фенологических времен года — естественных сезонов, подсезонов и этапов года в различных районах лесной части СССР

Продолжение при л. 4

Продолжение прил. 4

Окончание при л. 4

Приложение 5. Средний многолетний (1950-1982 гг.) феноспектр сезонного развития видов хвойных деревьев под Ленинградом
(условные обозначения см. на с. 322)

сплошная кривая- средняя температура воздуха (Т°С); пунктирная — продолжи-тельность светового дня (Сд), ч; точечная — влажный дефицит воздуха в 13 ч (Е, мбар)
1 — весенний рост (набухание) вегетативных почек; 2 — их распускание; 3 — рост и облиствение (охвоение) побегов; 4 — ассимиляционная деятельность сформированных листьев (хвои), опробковение и одревеснение побегов, формирование почек возобнов-ления; 5-осеннее расцвечивание и опадение листьев (хвои); б-многолетняя хвоя зимнезеленых хвойных пород; 7 — цветение (у хвойных — пыление и восприятие пыльцы семязачатками); 8 -формирование плодов (шишек) и семян; 9 — их созревание
1 — пихта сибирская; 2 — ель европейская; 3 — сосна обыкновенная; 4 — сосна кедро-вая сибирская; 5 — сосна кедровая корейская: 6 — лиственница Гмелина; 7 — листвен-ница сибирская; 8 — лиственница европейская; 9 — лиственница европейская поздно-распускающейся формы
Приложение 6. Средний многолетний (1950-1982 гг.) феноспектр сезонного развития видов лиственных деревьев под Ленинградом

Условные обозначения в феноспектре те же, что и в приложении 5. Виды лиственных деревьев:
Фенологическое развитие древесных растений
Под фенологическим развитием растений понимают закономерное чередование и ежегодное повторение одних и тех же фенологических циклов (вегетации и покоя, роста побегов и его прекращения, цветения, созревания плодов и семян и др.), а в пределах циклов — последовательный ход наступления и прохождения фенологических фаз роста и развития. Фенологическая фаза (фенофаза) — это такой этап в годичном цикле развития растения в целом или его отдельных органов, который характеризуется четко выраженными внешними морфологическими изменениями (появлением всходов, распусканием семядолей, набуханием и распусканием почек, развертыванием листьев, началом и окончанием роста побегов, цветением и созреванием плодов, расцвечиванием и опаданием листьев и др.). Календарное время наступления той или иной фенофазы называют фенодатой, а временной интервал между определенными фенодатами составляет межфазный период, или фенологический цикл.
В основе фенологического развития растений лежит наследственно закрепленная ритмичность и периодичность физиологических процессов, получившая название биологических, или физиологических, часов. Однако динамика наступления фенофаз, сроки начала, окончания и продолжительность фенологических циклов у растений находятся под постоянным и мощным воздействием сезонных изменений географической среды (при-роды) и прежде всего сезонности климатических условий (закономерного чередования на Земле сезонов с различной продолжительностью дня и ночи, теплых и холодных, дождливых и сухих), приспосабливаясь к которым, растения существенно изменяют ритмику процессов роста и развития, свое фенологическое состояние. В теплые или дождливые сезоны растения вегетируют, в холодные или сухие впадают в покой. Под влиянием сезонных изменений погодных условий у растений резко изменяется динамика их ростовых процессов. Поэтому фенологическое развитие растений понимают как их сезонное развитие.
Наука, синтезирующая в себе всю систему знаний о сезонном развитии природы, получила название фенологии. Ее раздел, изучающий сезонное развитие растений и образуемых ими сообществ, определяют как фитофенологию, а раздел о сезонном развитии древесных растений и их сообществ — как дендрофенологию. Фенология базируется на фенологических наблюдениях, в процессе которых устанавливают календарное время наступления изучаемых сезонных явлений, а у растений — фенофаз. Диапазон научных и прикладных задач, решаемых современной фенологией и дендрофенологией, чрезвычайно широк. В лесном хозяйстве, на-пример, по материалам многолетних фенологических наблюдений устанавливают закономерные связи между временем наступления тех или иных фенологических явлений-индикаторов и оптимальными сроками проведения работ по посеву и посадке леса, рубкам ухода, защите лесов от пожаров, вредных насекомых и болезней, по заготовке плодов, семян, ягод, грибов, лекарственного и дубильного сырья, по содействию естественному возобновлению леса. Фенологическое состояние лесов учитывают и при их таксации , (учете) с применением аэро- или космической фотосъемки, так как оптические свойства лесов тесно связаны с их сезонным развитием.
Велико значение фенологических наблюдений и в практике озеленения городов и населенных мест. Изучение динамики сезонного развития растений необходимо при подборе их для озеленения, для оценки эстетических и санитарно-гигиенических свойств растений, при разработке и проведении мероприятий, обеспечивающих повышение биологической устойчивости городских зеленых насаждений, их защиту от вредителей и болезней. Материалы фенонаблюдений используют при составлении календарей цветения растений, созревания и сбора плодов и семян, при установлении оптимальных сроков посева и посадки.
Многолетние данные о сроках наступления фенофаз (сокодвижения, распускания листьев, зацветания, созревания плодов, осеннего расцвечивания листьев и их опадания) у многих древесных пород (клена, березы, дуба, рябины, липы, ольхи и др.) широко используют при разработке систем естественной (фенологической) периодизации года (см. приложение 4) и для составления местных фенологических календарей. При изучении древесных растений фенологические наблюдения над ними дают обширную и ценную информацию о биологических особенностях растений и их экологических свойствах. С современным уровнем развития отечественной и зарубежной фенологии и дендрофенологии, их научным и практическим значением подробнее можно познакомиться по книгам Г. Э. Шульца (1980) и Н. Е. Булыгина (1982).
Сроки наступления различных фенофаз устанавливают в зависимости от целей фенологических наблюдений над древесными растениями. В одних случаях учитывают только отдельные фазы -индикаторы сезонного развития природы или наиболее хозяйственно важные, такие, как зацветание, созревание плодов и семян, их распространение. Если же нужно получить достаточно полное представление о фенологических особенностях растений того или иного вида (формы), наблюдениями охватывают все основные фе-нофазы, программа которых и условные обозначения фенофаз приведены ниже.
Программа фенологических наблюдений над древесными растениями
I. Растения на ювенильном этапе онтогенеза Развитие подсемядольной части растения: появление всходов (С); распускание семядолей (С2).
Развитие надсемядольной части растения: распускание (раскрывание) зародышевой почки (Пп1); распускание ювенильных листьев (Лп1); начало и оконча-ние роста осевого побега в длину (Пп2 и Пп3); опробковение побега по всей длине (Пи ); расцвечивание отмирающих ювенильный листьев (Лп2); опадение ювенильных листьев (Лп3).
II. Растения на виргинильном и последующих этапах онтогенеза
Наблюдения над вегетативными побегами. Растение в состоянии покоя; почки не имеют признаков роста (Пб°). Рост вегетативных материнских почек *: набухание почек (Пб1); распускание почек (Пб2).
Рост и формирование побегов продолжения: начало (Пб3) и окончание (Пб4) роста побегов в длину; опробковение побегов у основания (Пб5) и по всей длине (Пб6).
Формирование и отмирание листьев: распускание листьев (Л1); расцвечивание отмирающих листьев (Л3); опадение листьев (Л4).
Формирование почек возобновления: обособление на побегах почек в виде мелких зеленых бугорков (Пч1); опробковение с поверхности наружных чешуи почек (Пч2).
Наблюдения над генеративными и генеративно-ростовыми побегами **. Генеративные (генеративно-ростовые) почки находятся в состоянии покоя (почки не имеют признаков роста) — (Ц°).
Рост генеративных (генеративно-ростовых) почек: набухание почек (Ц1); распускание почек (Ц2).
Бутонизация и цветение***: бутонизация (Ц3); начало цветения (Ц4), окончание его (Ц5).
Формирование и созревание плодов (шишек) и семян: заложение плодов или шишек (Пл1); созревание плодов (шишек) и семян (Пл3).
Распространение зрелых семян: опадение зрелых плодов (шишек) с семенами или высыпание семян из плодов и шишек (Пл4 ).
* Материнскими называются почки, формирующиеся на растущих или закончивших рост побегах. Почки возобновления, которые закладываются (образуются) в материнских почвах, называют дочерними. После распускания материнской почки и вырастания из нее побега продолжения заложенные на этом побеге дочерние почки сами становятся материнскими.
** Генеративные побеги связаны только с образованием генеративных органов; закладываются они в функционально генеративных (цветковых) почках и после окончания цветения или созревания семян отмирают. Генеративно-ростовые побеги служат для образования как генеративных, так и вегетативных органов, они закладываются в генеративно-ростовых (смешанных) почках. После окончания цветения или созревания семян на побегах этого типа отмирает их генеративная часть, а вегетативная сфера сохраняется для выполнения функций вегетативных побегов.
*** У голосеменных растений фазе бутонизации соответствует фаза обособления на побегах микро- и макростробилов, а фазе цветения — фаза пыления. Последовательность прохождения фенофаз вегетативными и генеративными (генеративно-ростовыми) побегами в целом соответствует последовательности, указанной выше. Исключение составляют следующие случаи:
У большинства видов древесных растений распускание и рост листьев происходят параллельно с ростом побегов; у представителей рода сосна распускание молодой хвои начинается перед окончанием их роста.
Обособление и последующее формирование материнских почек происходят в период роста побегов, процессы формирования почек возобновления обычно завершаются после окончания роста побегов. У целого ряда древесных пород (кипариса, туи, платана, белой акации, актинидии) фенологические фазы формирования почек внешне не выражены.
Генеративные, а у таких родов древесных растений, как рябина, боярышник, калина, и генеративно-ростовые почки весной трогаются в рост раньше вегетативных.
Большим фенологическим разнообразием отличаются древесные растения по соотношению во времени прохождения циклов вегетативного и генеративного развития. Так, по периодам цветения различают древесные растения, цветущие до распускания листьев (осина, виды ольхи, лещины, вяза; до распускания молодой хвои пылит сосна), в начальный период облиствения (виды березы), вскоре после массового облиствения (дуб черешчатый, бук лесной, виды ореха), после окончания роста побегов — в середине лета (виды липы) или только в конце лета — осенью (аралия маньчжурская, целебник).
Особую биологическую группу составляют виды с ремонтантным цветением — очень продолжительным, циклически многократным. Такой тип цветения присущ растениям тропиков, но может наблюдаться и у ряда видов древесных растений умеренных широт (роза морщинистая, курильский чай). Иногда у древесных растений происходит вторичное цветение — преждевременное зацветание в конце лета или осенью цветков, заложенных в почках для будущего года (ива козья, конский каштан, слива, вишня, яблоня).
Значительно различаются между собой древесные растения разных родов и видов по времени созревания плодов и семян. Например, у осины и многих видов ивы семена созревают в конце весны; у вяза — в начале лета; у вишни, смородины, березы повислой — в середине лета; у рябины — в конце. Плоды дуба черешчатого и бука, семена сосны обыкновенной и кедровой сибирской, пихты, ели созревают в начале осени; плоды кленов остро-листного и ложноплатанового, клюквы — в середине осени, а липы и ольхи — в конце. Еще более различны у древесных растений разных фенологических групп периоды распространения зрелых плодов и семян.
Далеко не одинаковы у древесных растений и циклы формирования плодов и семян. У ивы, тополя и вяза их созревание происходит через 3-6 недель после зацветания; у дуба черешчатого и скального — через 3-4 месяца; у сосны, кипариса и дуба из секции красных дубов (дуб красный, шарлаховый и др.) — только через 1,5 года.
В связи с изменениями климатических и других условий внешней среды у одних и тех же видов и форм древесных растений одноименные фенофазы могут наступать в совершенно различные сроки как в одном географическом пункте, так и в разных. Поэтому регулярные многолетние фенологические наблюдения очень важны для установления фактических местных периодов наступления фенофаз у изучаемых растений. Эти же фенологические наблюдения позволяют выявлять истинную длительность разного рода фе-нологических циклов, прежде всего циклов вегетации и покоя.
Вегетация — это такое состояние растений, при котором происходят процессы видимого роста вегетативных и генеративных органов и осуществляется непрерывная ассимиляционная деятельность листьев.
Покой — период (во временном отношении — цикл) в годичном цикле развития растений, когда видимый рост отсутствует, а листья не ассимилируют, в том числе и у вечнозеленых растений.
У листопадных видов древесных растений фенологическим индикатором начала вегетации является распускание вегетативных почек, а окончания ее — полное осеннее расцвечивание листьев в кроне или их опадание, если листья опадают зелеными (сирень обыкновенная, жостер слабительный, ольха серая, часто ясень обыкновенный). У вечнозеленых видов фенологические признаки начала и окончания вегетации менее определенны.
Растения со сходными сроками начала и окончания вегетации и близкой продолжительностью циклов вегетации и покоя объединяют в определенные фенологические группы, получившие название феноритмотипа. Примерами различных феноритмотипов древесных растений могут служить следующие. Во влажно-тропических лесах феноритмотип составляют вечнозеленые, непрерывно вегетирующие деревья и кустарники. У них не бывает периодов покоя, а наблюдается чередование циклов роста побегов и временной приостановки его. Иной феноритмотип составляют вечнозеленые древесные растения умеренного климатического пояса: они вегетируют летом, зимой находятся в покое, весенний сезон для них является периодом перехода от состояния покоя к вегетации, а осенний — от вегетации к покою.
Свои феноритмотипы имеют листопадные растения. Одни из них характеризуются ежегодным чередованием одного цикла вегетации и одного цикла покоя (например, древесные породы мелко- и широколиственных лесов СССР), другие же могут вегетировать и впадать в покой (вынужденный) несколько раз за год. Так, по свидетельству Г. Вальтера (1968), в субтропической пустыне Соноре (Северная Америка) есть виды кустарников, вегетирующих до 10 раз в году и столько же раз вступающих в покой. Такая многоцикличность вегетации и покоя обусловлена периодичностью выпадения дождей: как только почва достаточно увлажнится, растения начинают вегетировать, а с прекращением дождей и наступлением очередного засушливого периода они сразу сбрасывают листья и впадают в покой.
Характерные особенности сезонного развития различных видов и фенологических форм хвойных и лиственных древесных растений СССР наглядно показаны на помещенных в приложениях 5 и 6 феноспектрах — графических изображениях последовательности наступления и прохождения растением фенологических фаз и циклов сезонного развития.
Фенологические наблюдения над древесными растениями (дендрофенонаблюдения) требуют четкого знания методики их проведения и особенно диагностических (морфолого-биологиче-ских) признаков наступления регистрируемых фенофаз. С этими признаками, условными обозначениями фенофаз, методикой проведения наблюдений и оформления их результатов рекомендуется знакомиться по специальным фенологическим руководствам (Булыгин Н. Е., 1974, 1976, 1979; Александрова Н. М. и др., 1975; Елагин И. Н., Лобанов А. И., 1979; Зайцев Г. Н., 1981; Булыгин Н. Е., Щеголева С. В., 1982). Образец отчетного бланка учебных дендрофенологических наблюдений приведен в приложении 7.
Современная дендрофенология изучает не только динамику внешне выраженных фенологических (макрофенологических) процессов, но и ритмику формирования зачаточных побегов в почках, образования годичного кольца древесины, заложения и формирования зачаточных органов семени. Весь этот круг исследований, выполняемых с использованием бинокулярных луп и микроскопов, относят к микрофенологии (Булыгин Н. Е., 1982), непосредственно смыкающейся с задачами изучения морфогенеза растений (морфогенез — процесс формообразования, т. е. заложения, роста и развития клеток, тканей, органов).
Существуют также специальные программы и методики, связанные с изучением динамики сезонного роста корней древесных растений. С этой специфической областью дендрофенологии можно ознакомиться по руководствам В. А. Колесникова (1972) и Г. Д. Ярославцева (1973).
1. Что понимают под «жизненной формой растений»? Какие жизненные формы относят к древесным и полудревесным растениям? Каковы морфобиологические особенности деревьев лесного, кустовидного, плодового, сезонно-суккулентного и стланцевого типов?
2. Какие жизненные формы растений относят к кустарникам, полукустарникам, кустарничкам, древовидным, кустарниковым и полукустарниковым лианам, древесным растениям-подушкам?
3. Каковы морфобиологические особенности основных этапов онтогенеза древесных растений: эмбрионального, ювенильного, виргинильного, генеративного старения?
4. Что изучают науки «фенология» и «дендрофенология»? Каковы содержание; задачи, методы и значение дендрофенологии для практики лесного хозяйства и озеленения?
5. Какие процессы понимают под фенологическим развитием древесных растений, их фенологическим биоритмом, физиологическими, или биологическими часами, циклами вегетации и покоя, циклами вегетативного и генеративного развития?
6. Каковы основные фенологические фазы древесных растений, последовательность их прохождения у видов различных фенологических групп?
7. По наступлению и завершению каких фенофаз судят о сроках начала и окончания вегетации у древесных растений? Как можно объяснить понятие «феноритмотип» растений?
8. Какие вы знаете основные дендрофенологические индикаторы, по которым можно судить о фактических сроках наступления естественных сезонов и подсезонов года в различных районах лесной части СССР?
9. Каковы внешние факторы и практическое значение ежегодной и географической изменчивости периодов наступления и прохождения фенофаз у растений и фенологических времен года?